Coral Reefs worldwide are at risk of collapse by climate change. This will result in devastating impacts to marine fisheries and biodiversity, to planetary-level ecosystem services such as carbon sequestration, and to the livelihood of over 500 million people who depend on coral reefs. The coral host depends on the photosynthetic carbon translocated from its algae endosymbiont partner, but this relationship can breakdown (coral bleaching) under continuous temperatures of only 1-2° C above the average monthly historical maxima and lead to coral mortality. However, coral species show differential bleaching response, with some bleaching and dying and others resisting bleaching or quickly recovering.
Marcelino's Lab at Northwestern University
Climate change effects on coral reefs, coral adaptation and acclimatization to warmer oceans
RESEARCH PROJECTS
Identifying successful coral-algal partnerships for heat resistance
Swain et al., 2021 mSystems
Swain et al., 2017 FunctEcol
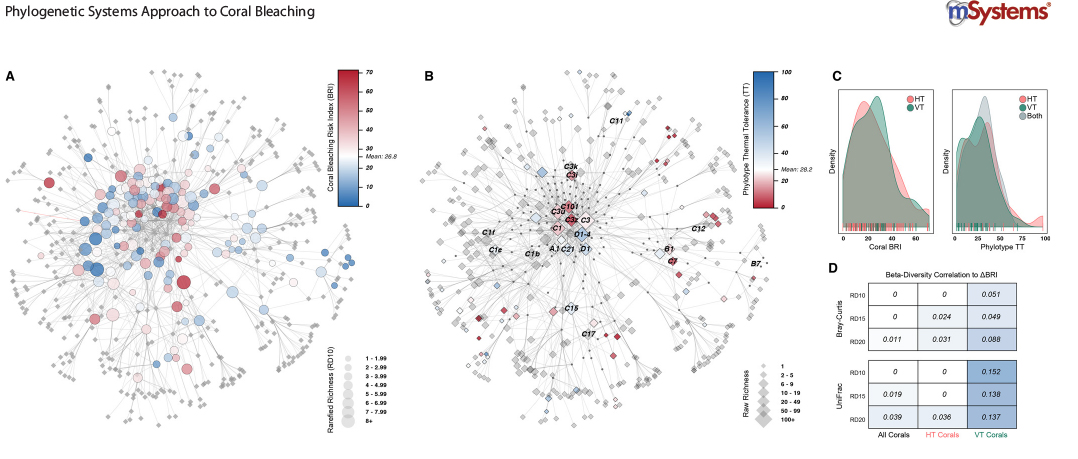
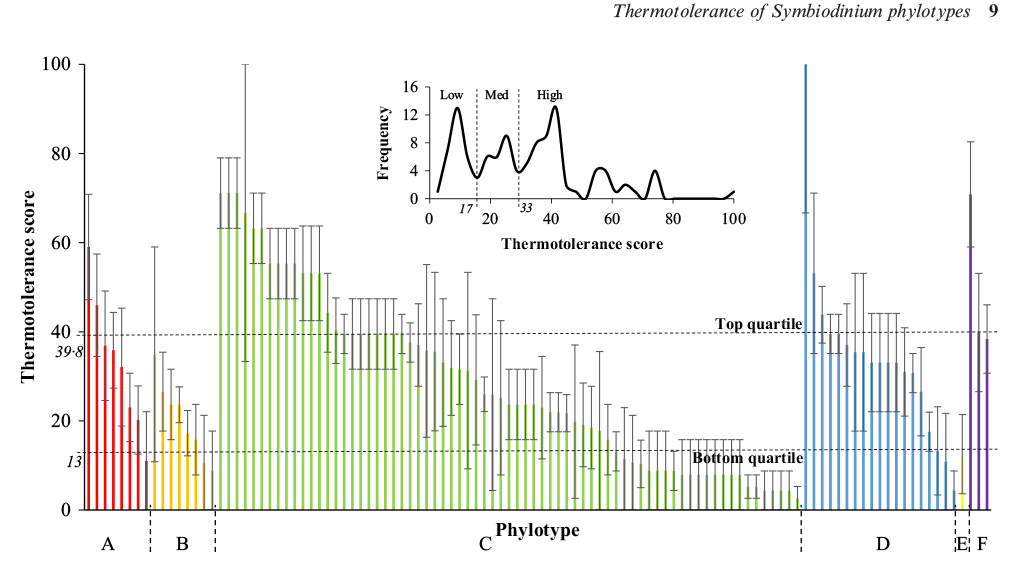
We are developing algorithms and using cluster analysis, neural network analysis, and phylogenetically-corrected statistical analysis to identify the coral-algal relationships that have a higher likelihood to survive heat stress episodes. We have built a coral bleaching and mortality index, an algal tolerance to stress index, and coral-algal association networks to identify whether corals that associate with thermal tolerant algae show increased resistance to heat.
Our goal is to uncover biological factors in both partners that promote a successful response to heat stress and to identify the most heat-tolerant genetic combination of algal-coral partners to build more heat resistant coral stocks (adaptation through “assisted evolution”).
Relevant Publications
Swain, T. D., Lax, S., Gilbert, J., Backman, V. and Marcelino, L. A., 2021, A phylogeny-informed analysis of the global coral-Symbiodiniaceae interaction network reveals that traits correlated with thermal bleaching are specific to symbiont transmission mode. mSystems. 6, 3, e00266. PMCID: PMC8269218
Swain, T. D., Lax, S., Backman, V., and Marcelino, L. A. 2020, Uncovering the role of Symbiodiniaceae assemblage composition and abundance in coral bleaching response by minimizing sampling and evolutionary biases, BMC Microbiology 20, 1, 124, PMCID: PMC7236918
Swain T.D., Bold E.C., Osborn P.C., Baird. A.H., Westneat M.W., Backman V., Marcelino L.A. 2018, Physiological integration of coral colonies is correlated with bleaching resistance, Mar Ecol Prog Ser, 586: 1–10, [Feature Article] DOI
Swain TD, Westneat WM, Backman V, Marcelino L.A. 2018, Phylogenetic analysis of symbiont transmission mechanisms reveal evolutionary patterns in thermotolerance and host specificity that enhance bleaching resistance among vertically transmitted Symbiodinium. European Journal of Phycology 53, 1-17. DOI
Swain, T.D., DuBois, E., Goldberg, S.J., Backman V., Marcelino, L.A., 2017, Bleaching response of coral species in the context of assemblage response, Coral Reefs 36: 395–400. PMCID: PMC5679474
Swain T.D, Vega-Perkins J.B., Oestreich W., Triebold C., DuBois E., Henss J., Baird A, Siple M., Backman V., Marcelino L.A. 2016, Coral bleaching response index: a new tool to standardize and compare susceptibility to thermal bleaching, Glob Chang Biol. PMCID: PMC5433437
Swain T.D, Chandler J.*, Backman V., Marcelino L.A., 2016, Consensus thermotolerance ranking for 110 Symbiodinium phylotypes: an exemplar utilization of a novel iterative partial rank aggregation tool with broad application potential. Functional Ecology, DOI
Understanding the role of light in coral physiology and bleaching susceptibility
Marcelino et al., 2013 PLoS1

Su Y. et al., manuscript in preparation
Spicer et al., 2019 Scientific Reports
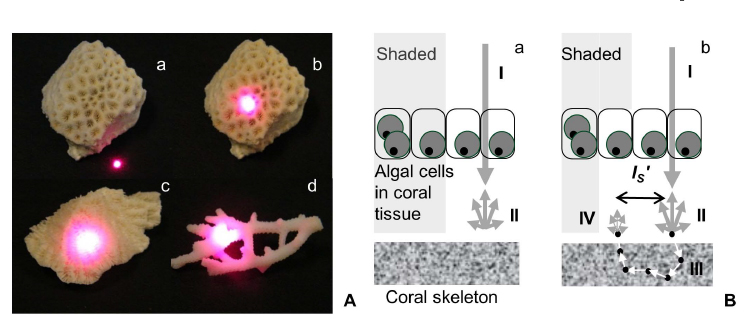
Corals are like greenhouses growing millions of symbiont algae inside their cells which provide the coral with more than 90% of its daily energy requirement. The coral can be viewed as an intricate optical machine and its interaction with visible light is one of the key factors in the coral life cycle and bleaching episodes. The coral consists of two compartments: 1) the living tissue where the host photoprotective pigments and the symbiont algae with all its photosynthetic pigments are contained, and 2) the skeleton, a highly reflective limestone structure secreted by the coral polyp in a species-specific manner and affected by local environmental factors.
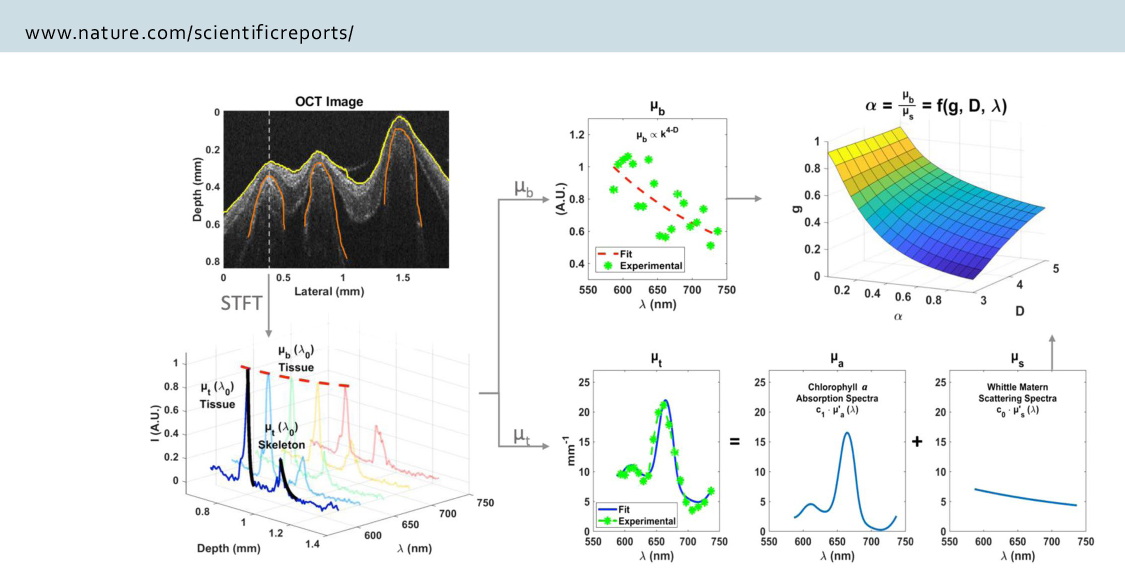
Coral skeletons are highly scattering in the upper ~200 μm of the skeleton layer (measured as ‘microscopic’ reduced coefficient, µSʹ,m ) and effectively amplify and homogenize the ambient light-field of their endosymbiotic dinoflagellates. This effect enhances productivity of the symbiosis, but also contributes to the likelihood of catastrophic dissociation as densities of light-absorbing symbionts decrease and light levels for the remaining symbionts rapidly increase.
We are studying light transport and scattering in coral to better understand its role on coral physiology and coral bleaching susceptibility. We are using photophysiological techniques, light scattering models and experiments, and non-invasive imaging techniques such as Inverse Spectroscopic Optical Coherence Tomography (ISOCT).
Relevant Publications
Spicer G.L.C., Eid A., Wangpraseurt D., Swain T.D., Winkelmann J.A., Yi J., Kühl, Marcelino L.A., Backman V., 2019, Measuring light scattering and absorption in corals with Inverse Spectroscopic Optical Coherence Tomography (ISOCT): a new tool for non-invasive monitoring, Scientific Reports 9:14148 PMCID: PMC6775107
Swain T.D., Lax S., Lake N., Grooms H., Backman V., Marcelino L.A., 2019, Relating coral skeletal structures at different length scales to growth, light availability to Symbiodinium, and thermal bleaching, Frontiers in Marine Science 586 1-10 PMCID: PMC4800776
Swain T.D., DuBois E., Gomes, A., Stoyneva V.P., Radosevich A.J., Henss J., Wagner M.E., Velazquez E., Traub J., Kennedy B.J., Janczak C.M., Grigorescu A.A., Westneat M.W., Sanborn K., Levine S., Schick M., Parsons G., Rogers J.D., Backman V., Marcelino L.A. 2016, Skeletal light scattering decreases bleaching susceptibility of reef-building corals, BMC Ecol. 16(1):10. PMCID: PMC4800776
Swain TD, Westneat WM, Backman V, Marcelino L.A. 2018, Phylogenetic analysis of symbiont transmission mechanisms reveal evolutionary patterns in thermotolerance and host specificity that enhance bleaching resistance among vertically transmitted Symbiodinium. European Journal of Phycology 53, 1-17. DOI
Marcelino L.A., Westneat M., Stoyneva V.*, Henss J., Rogers J.D., Radosevich A.*, Turzhitsky V.*, Siple M.*, Fang A.*, Swain T.D., Fung J.* and Backman V. U 2013, Light scattering in coral skeleton - evolutionary trends in coral bleaching and in light amplification, PLoS One, 8 :e61492. PMCID: PMC3632607
Su Y., Wangpraseurt D., Eid A., Spicer G.L.C., Winkelmann J.A., Swain T.D., Yi J., Kühl M., V. Backman and L. A. Marcelino, Light scattering, absorption properties of tissue and coral skeleton and photophysiology of 4 scleractinian corals using Inverse Spectroscopic Optical Coherence Tomography (ISOCT) and Pulse amplitude modulated (PAM) fluorometry, in preparation
Increasing Coral’s Adaptive Response to Warmer Oceans
.jpg)
Heat-stressed (white fragments, front) and Control (brown fragments, back) Acropora cervicornis (Florida Keys, in collaboration with Dr. Ross Cunning and Dr. Shayle Matsuda at The Shedd Aquarium).
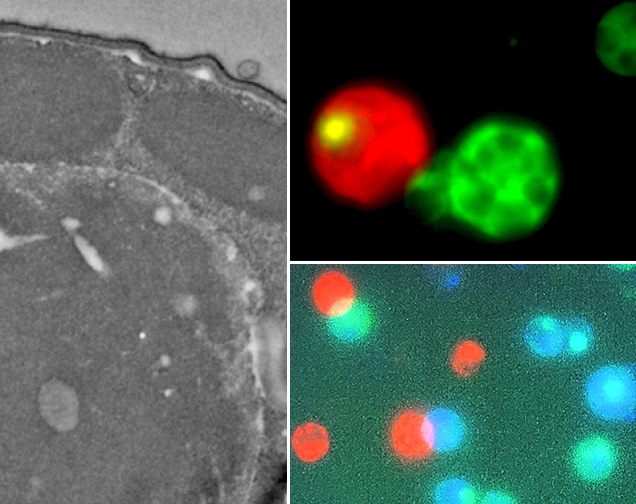
Fluorescence microscopy and Electron microscopy in different types of coral cells
Total RNA quality analysis for RNA-seq
We are studying the epigenetic processes underpinning a successful response to heat stress in corals that increases their likelihood of survival. Specifically, we are measuring differences in gene expression between heat-resistant and heat-sensitive corals and uncovering differences in transcription regulation mechanisms between these corals, such as epigenetic marks (e.g., acetylation and methylation) and changes in structure of chromatin (DNA and protein complex). Our main goal is to boost coral adaptation and resilience to a fast-warming ocean via rapid physiological changes to heat stress that increase their likelihood of survival. These processes are driven by genetic differences (“thermal genes”) and changes in gene expression that do not involve alterations in the genetic code (epigenetics).
We use a combination of techniques such as Chromatin sequencing (ChIP-seq and ATAC-seq), Transcriptomic sequencing (mRNA-seq), Immunohistochemistry, Chromatin imaging (Chromatin Electron Microscopy, ChromEM and Partial Wavelength Spectroscopy, PWS) in collaboration with the Center for Physical Genomics and Engineering at Northwestern University and the Shedd Aquarium in Chicago.
Relevant Publications
Eid A., Worsfold, F., Backman V., and Marcelino L.A. Modeling the differential adaptative resistance to bleaching in corals – bleaching and recovery of Florida Reef Track corals 2005 - 2019, in preparation
Li W.S., Acosta N., Shen K., Cunning R., Matsuda S., Bleher R., Backman V., and Marcelino L.A., Chromatin modifications are key regulators in gene expression reprograming during coral bleaching resistance, in preparation